第三节 载脂蛋白基因的多态性
在人类进化过程中,基因的DNA序列在逐渐演变。这种演变大都发生在氨基酸编码子以外的区域,称为“中性”变化,不导致临床疾病的发生。但是,基因的氨基酸编码区以及基因表达的调控区的DNA序列改变时有发生。导致蛋白质的合成量改变或基功能的异常。载脂蛋白结构基因或其调控区DNA序列的突变引起致高脂血症或早发动脉粥样硬化已大量见诸于医学文献。新的载脂蛋白基因的多态性不断地被发现。
检测载脂蛋白基因的遗传变异通常使用从外周血白细胞中提取的DNA为起始材料,通过聚合酶链反应扩增基因DNA的产量,嗣后,或采用变性梯度凝胶电泳(denaturing gradientgel electrophoresis, DGGE)显示由基因编码胺基酸区DAN序列突变所致的单股DNA构象的多态性;或使用等位基因特异的寡核苷酸探针杂交法(allele-specificoligonuc leotide hybridization,ASO)探测突变的等位基因;或测定限制性片段长度多态性(restriction fragment length polymorphism, RFLP)鉴定基因DNA序列的突变;或直接对某基因的片段进行DNA序列测定,与正常的相应的DNA序列比较,检测有否突变发生。这几种方法为较常使用的方法。有关这些方法的详细介绍可参见本书第十六章的生物化学检验技术篇。本节主要介绍ApoB,ApoE,Apo(a)。ApoAⅠ基因的多态性。对其他载脂蛋白基因的多态性仅以举例的方式略述。
一、ApoB基因的遗传变异与多态性ApoB基因的遗传变异与多态性分以下几种类型进行介绍:①RFLP;②导致血浆胆固醇水平降低的基因突变;③导致血浆胆固醇升高的基因突变;④其他类型的多态性。
1.RFLP
据报道,载脂蛋白B基因的限制性片段长度多态性至少375种。后来发现有些多态性并非存在,纯属DNA序列测定错误。表4-3列举了10种RFLP。产生这些RFLP的突变或发生在ApoB基因上游启动子区域,或发生在内含子区域,或在外显子区域。
表4-3 ApoB基因限制性片段长度多态性举例
| 惹限制性内切酶 | 酶切位点 | 突变等位基因出现频率 | 酶切部位存在或缺失 |
| AvaⅡ | 距第1外显子上游4Kb | 0.20 | 缺失 |
| MspⅠ | 启动子区内,-265bp | 0.21 | 缺失 |
| ApaLⅠ | 第4外显子,cDNA的417bp | 0.36 | 缺失 |
| HincⅡ | 第4内含子内,距第3外显子3‘末端3334bp | 0.12 | 存在 |
| PvuⅡ | 第4内含子内,距5外显子5‘末端4523bp | 0.08 | 存在 |
| AluⅠ | 第14外显子内,cDNA的1981bp | 0.48 | 缺失 |
| BalⅠ | 第20内含子内,距第21外显子5‘末端146bp | 0.50 | |
| XbaⅠ | 第26外显子内,cDNA的7674bp | 0.40-0.50 | 存在 |
| MspⅠ | 第26外显子内,cDNA的11040bp | 0.12 | 缺失 |
| EcoRⅠ | 第26外显子内,cDNA的12670bp | 0.20 | 缺失 |
(源自:Young S G. Circulation 82:1583,1990)
表4-3所列发生在外显子内的突变,除XbaⅠ限制性片段长度多态性外,其余均导致氨基酸改变。已有很多研究者致力于研究常见的ApoB基因PFLP与高胆醇血症、高甘油三酯血症硬化的相关关系,但发现令人信服的相关关系的报道不是很多,还有的结果相互矛盾。正如Young指出的那样,有的研究所使用的样本量太小;有的研究对照组的选择不合规范;有的研究基于不同的种族,忽视了正常的种族差异。很显然。在以后的相关研究中,尽可能选用大样本量,合理的选择对照组,考虑不同种族的正常遗传变异无疑对确证某些ApoE基因的RFLP与脂蛋白代谢紊乱的特殊表型以及早发动脉粥样硬化的关联尤为重要。
2.导致血浆胆固醇水平降低的ApoB基因突变
经过许多实验室的努力,一些ApoB基因的无义突变或移码突变导致家族性密度脂蛋白血症均已鉴定(见表4-4)。
表4-4 ApoB基因突变导致短ApoB的合成
| ApoB的长度 | DNA水平上的突变 | 短ApoB在血浆脂蛋白中的分布 |
| B-2 | 在第5内含因子中,G颠换成T | 无 |
| B-9 | T替代C,使翻译在411号密码子后终止 | 无 |
| B-25 | 在第21外显子内有694bp缺失 | 无 |
| B-27.6 | 在第24内含子内,剪接部位突变 | HDL,LDL |
| B-29 | 在第4152核苷酸上,T替代C | 无 |
| B-31 | cDNA核苷酸4480缺失 | HDL及密度大于1.2g/cm3部分 |
| B-32 | 在密码子1450上,T替代C | HDL,LDL |
| B-32.5 | 在4631核苷酸上,G替代T | HDL及密度大于1.21g/cm3部分 |
| B-37 | cDNA核苷酸5391至5394段缺失 | VLDL,LDL,HDL |
| B-39 | cDNA核苷酸5591缺失 | VLDL,LDL |
| B-40 | cDNA核苷酸5693至5694段缺失 | VLDL,LDL,HDL |
| B-46 | 在cDNA核苷酸6381上,T替代C | VLDL,LDL,HDL |
| B-49.6 | 在cDNA核苷酸6963上,T替代C | VLDL,LDL,HDL |
| B-52.8 | cDNA核苷酸7295缺失 | VLDL,HDL |
| B-54.8 | cDNA核甘酸7539缺失在cDNA核苷酸7655上,T替代C | VLDL,HDL |
| B-61 | 在核苷酸8525处,有37bp缺失 | VLDL,HDL |
| B-67 | cDNA核苷酸9327缺失 | LDL,VLDL |
| B-74.7 | cDNA核苷酸10366缺失 | LDL,VLDL |
| B-82 | 在核苷酸11411上,A替代C | VLDL,HDL |
| B-86 | cDNA核苷酸11840缺失 | VLDL,HDL |
| B-87 | cDNA核苷酸12032缺失 | VLDL,HDL |
| B-89 | cDNA核苷酸12309缺失 | VLDL,HDL |
(源自:Kane ,J.P.,and Havel, R.J.In:The metalbolic and molecular bases of inherited disease,edited by Scriver, C.R.et al, Vol Ⅱ,p1868,1995,McGraw –Hill,Inc.,New York,U.S.A)
家族性低低密度脂蛋白血症的患者大多为杂合子,即他们携带一个正常的ApoB等位基因和一个变异的ApoB等位基因。纯合子以及复合杂合子患者少见。短ApoB合成的患者血浆中ApoB的浓度较低,仅为正常人ApoB等位基因产生的ApoB-100浓度的2%~10%,其机理尚未完全明了。有人发现,含有ApoB87和ApoB89的LDL比含ApoB-100的LDL更易于被LDL受体摄取。亦有人推测在杂合子患者中,由一个正常ApoB-100等位基因编码的ApoB-100与两个正常ApoE等位基因编码的ApoE组成的脂蛋白颗粒比正常的脂蛋白颗粒多含有1倍的ApoE,故前者易于被LDL摄取清除。这些均有助于解释一部分短ApoB合成患者体内低浓度血浆ApoB及低浓度血浆胆固醇的机制有等进一步努力。不难想到的是,家族性低低密度脂蛋白血症患者血浆中LDL胆固醇低,产生冠心病及动脉粥样硬化的危险性也降低。已有一些报道证实,这些患者不产生动脉粥样硬化且寿命增长。故有人认为,对惯常食用高饱和脂肪、高胆固醇饮食的北美人群,携带合成短ApoB的突变基因非但无害,反而有益于健康。
3.导致血浆胆固醇升高的基因突变。
Brown和Goldstein的经典研究,即LDL受体基因的突变,引起LDL清除障碍,LDL在血浆中蓄积,导致早发动脉粥样硬化,这是家族性高胆固醇血症的主要病因。这些患者的血浆总胆固醇浓度高于7.76mmol/L。然而有相当一部分人的血浆胆固醇浓度在6.47~7.76mmol/L之间,与正常人相比为轻中度升高。长期以来,人们一直推测ApoB基因遗传变异可合成有缺损的ApoB蛋白质,后者与LDL受体亲和力下降,引起LDL清除障碍,LDL在血浆中蓄积。这种推测于1989年得到了证实。Soria等人发现了家族性ApoB100缺损。这些病人在其Apob cDNA第10708个核苷酸位置上,G转换成了A,导致ApoB100的等3500个氨基酸精氨酸变换成了谷氨酰胺。这一突变使其LDL与LDL受体的结合力下降,产生高胆固醇血症。据估计,这种突变在人群中的发生率与LDL受体的突变率接近,即1/500。
另一家族性ApoB100缺损是由ApoB基因第10800核苷酸位置上的C转换成T所致。这一突变ApoB内第3531个氨基酸精氨酸变成了半胱胺酸。8个患者的平均血浆LDL胆固醇水平在6.78mmol/L,而8个未患ApoB基因突变的亲戚为3.88mmol/L。患者LDL对LDL受体的亲和力仅为正常对照组的39%。由于C至T的转换,产生一个新的限制性内切酶切割部位(NsiI),这将有助于在大范围人群内检测这种突变发生率。
4.其他类型的多态型
距ApoB基因3‘末端500个核苷酸碱基对区域,有一DNA片段构成可变动数目的半联重复(variable number of tandem repeats. VNTR)已发现ApoB基因3‘末端这种VNTR所致的多态性与心肌梗塞的发作有关联。
ApoB信号肽由27个氨基酸组成。在高加索种族人群中发现了两种多态性:一种为插入等位基因型,编码27个氨基酸;另一种为缺失等位基因型,编码24个氨基酸。Visvikis 等人发现缺失等位基因型与高血浆ApoB 水平、LDL胆固醇水平、LP(a)水平以及总胆固醇水平有关。最近Kammerer等人在34家共686名美籍墨西哥人中发现编码ApoB信号肽的三种等位基因:SP-24,SP-27,SP-29,它们分别编码24,27与29肽。携带SP-24等位基因的纯合子其血浆ApoB与LDL胆固醇的浓度显著高于SP-27纯合子。SP-29杂合子的ApoB与LDL胆固醇水平比SP-24纯合子还高。
二、ApoE基因的多态性ApoE在胆固醇与甘油三酯代谢中起着重要作用。在正常人群中,10%以上的个体间血浆胆固醇水平的着异可归究于ApoE基因的多态性。主要的ApoE等位基因有ApoE2、ApoE3和ApoE4。ApoE3在人群中的出现率最高,被认为较常见的ApoE基因。ApoE2与Ⅲ型高脂蛋白血症相关,而ApoE4在老年性痴呆病(Alzheimer‘s disease)患者中的出现率较高。这后一发现开拓了人们的视野,即ApoE不仅在脂类代谢中至关重要,而且在其他的代谢过程中也扮演重要角色。已有报道,ApoE参与组织损伤的修复、免疫调节以及调控细胞生长与分化。相信随着时间推移,更多新的ApoE功能及新的ApoE基因多态性会被发现。
表4-5列举了一些已鉴定的ApoE基因的遗传变异。表中同时列出了密码子及其编码的氨基酸的改变、特定ApoE基因变异型首先被鉴定出的那个民族、出现频率、相关的高脂血症型以及遗传方式。
表4-5 ApoE多态性
| 多态性名称1 | 变异密码 子 | 核苷酸改变 | 氨基酸改变2 | 种族背景 | 分布率3 | 高脂血症型4 | 遗传方式 | |
| 常见的ApoE基因型 | ||||||||
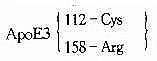 | - | - | - | 所有民族 | 41.9~91.1% | - | ||
| ApoE4 | 112 | TGC→CGC | Cys→Arg | 所有民族 | 6.4%~36.8% | N/HC | ||
| ApoE2 | 158 | CGC→TGC | Arg→Cys | 大部分民族 | 0.0%~14.5% | N/FD | 陷性 | |
| 与FD相关的多态型 | ||||||||
| ApoE0 | 第3内含子 | A →G,G缺 | 合成两种短肽 | 美洲黑人 | 14 | FD | 陷性 | |
| 3592bp-60 | 失移码突变 | 合成60肽 | 德国人 | 3 | FD | 陷性 | ||
| ApoE4-费城 | 13 | GAG →AAG | Glu →Lys | 拉丁美洲人 | 7 | FD | 陷性 | |
| 145 | CGT →TGT | Arg →Cys | 拉丁美洲人 | 陷性 | ||||
| ApoE3-Leiden | 112 | TGC →CGC | Cys →Arg | 荷兰人 | >40 | FD | 显性 | |
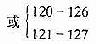 | 21bp重复 | 7个氨基酸患联重复 | ||||||
| ApoE1 | 127 | GGC→GAC | Gly→Asp | 高加索人 | >25 | FD | 未知 | |
| 158 | CGC→TGC | Arg→Cys | ||||||
| ApoE2-Christchurch | 136 | CGC→AGC | Arg→Ser | 未报道 | 3 | FD | 未知 | |
| ApoE3 | 112 | TGC→CGC | Cys→Arg | 拉丁美洲人 | 6 | FD | 显性 | |
| 142 | CGC→TGC | Arg→Cys | ||||||
| ApoE3--kochi | 145 | CGT→TGT | Arg→His | 日本人 | 4 | FD | 未知 | |
| ApoE2 | 145 | CGT→TGT | Arg→His | 未报道 | 3 | FD | 未知 | |
| ApoE1- Harrisburg | 146 | AAG→GAG | Lys→Glu | 日本人高加索 人 | 8 | FD | 未知 | |
| ApoE2 | 146 | AAG→GAG | Lys→Gln | 荷兰人美国人 | >40 | FD | 显性 | |
| ApoE3华盛顿 | 210 | TGG→TAG | Trp→终止密码 | 美国白人 | 3 | FD | 陷性 | |
| ApoE2 Fukuoka | 224 | CGG→CAG | Arg→Gln | 日本人 | 1 | FD | 未知 | |
| 与非FD高脂血症相关的多态型 | ||||||||
| ApoE5 | 3 | GAG→AAG | Glu→Lys | 日本人 | 0.5% | HC | ||
| 13 | GAG→AAG | Glu→Lys | 法籍加拿大人 | 6 | HC/HTG | |||
| ApoE2-Dunedin | 228 | CGC→TGC | Arg→Cys | 未报道 | 2 | HTG | ||
| ApoE2 | 236 | GTG→GAG | Val→Glu | 荷兰人 | 8 | HTG | ||
| ApoE7-Suita | 244 | GAG→AAG | Glu→Lys | 日本人 | 0.5%~0.8% | HC/HTG | ||
| 245 | GAG→AAG | Glu→Cys | ||||||
| ApoE3 | 112 | TGC→CGC | Cys→Arg | 荷兰人 | 3 | HTG | ||
| 251 | CGC→GGC | Arg→Gly | ||||||
| ApoE1 | 158 | CGC→TGC | Arg→Cys | 荷兰人 | 2 | HC | ||
| 252 | CTG→GAG | Leu→Glu | ||||||
| 与高脂血症无关的多态型 | ||||||||
| ApoE4-+-Ferburg | 28 | CTG→CCG | Leu→Pro | 德国高加索人 | 0.4%~0.9% | N | ||
| 112 | TGC→CGC | Cys→Arg | ||||||
| ApoE4 | 112 | TGC→CGC | Cys→Arg | 荷兰人 | 5 | N | ||
| 274 | CGC→CAC | Arg→His | ||||||
| ApoE4+ | 296 | AGC→CGC | Ser→Arg | 荷兰人 | 5 | N | ||
| ApoE5 | 84 | CCG→CGG | Pro→Arg | 美国白人 | 0.2% | N | ||
| 112 | TGC→CGC | Cys→Arg | ||||||
| ApoE3-Freibaug | 42 | ACA→GCA | Thr→Ala | 德国高加索人 | 1 | N | ||
| ApoE3 | 99 | GCG→ACG | Ala→Thr | 美国人 | 1 | N | ||
| 152 | GCC→CCC | Ara→Pro | ||||||
| ApoE2 | 134 | CGG→CAG | Arg→Gln | 荷兰人 | 5 | N | ||
1.除费城(Philadelphia)与华盛顿(Washington)两地名外,其他的地名均未翻译过来;
2.氨基酸代号:Cys-半胱氨酸;Arg-精氨酸;Glu-谷氨酸;Lys-赖氨酸;Gly-甘氨酸;Asp-天门冬氨酸;Ser-丝氨酸;His-组氨酸;Gln-谷氨酰胺;Trp-色氨酸;Val-缬氨酸;Leu-亮氨酸;Thr-苏氨酸;Ala-丙氨酸;Pro-脯氨酸;
3:分布率:加百分号的分布率是根据检测1000人以上的结果计算的;未加百分号的数值代表测出的携带ApoE多态型(包括纯合子与杂合子)的人数;
4:缩写:N(Normal,正常)HC(hypercholesterolemia,高胆固醇血症);FD(familialdysbetalipoproteinemia,家族性异常低密度酯蛋白血症,含Ⅲ型高脂蛋白血症)。
(源自:Knijff et al. Hum. Mutation4:181,1994)
三、Apo(a)基因的多态性血浆Lp(a)的浓度与Apo(a)蛋白质的大小成反比,即Apo(a)的分子量愈大,则Lp(a)的浓度愈低;Apo(a)的分子量愈小,则Lp(a)的浓度愈高。Apo(a)的分子量在400~800KDa之间。按照Apo(a)在聚丙烯凝胶电脉时相对于ApoB100(分子量为520KDa)的迁移率,Utermann等人于1987年将Apo(a)归类为六种多态型;F比ApoB100迁移快;B与ApoB100迁移率相等;S1,S2,S3,S4比ApoB100迁移慢。现已证实。Apo(a)蛋白质的多态型是由Apo(a)基因中Kringle-4的数目变化而引起的。Kring-4基因片段含有几个稀少的限制性酶切部位(NotI,SfiI.KspI,Sval,KpnI),这为加速筛选Apo(a)基因的多态性提供了帮助。表4-5列举在奥地利北部Tyrolean区高加索人群中用KpnI作工具所测得的RFLP与1-4Kringe的数目、Apo(a)蛋白质的大小以及血浆Lp(a)浓度的关系。
表4-6 Apo(a)Kpni 片段与Apo(a)蛋白质多态型、Kringle-4功能区数目以及血浆Lp(a)浓度之关系
| 表现分子量(KDa) | KpnI片段(Kb) | Kringle-4数目 | 等位基因 数目 | 平均Lp(a)浓度(mg%) | KpnI片段频率(%) | |
| F | <450 | 37-49 | 11-13 | 1-3 | * | <0.2 |
| B | ~500 | 55-66 | 14-16 | 4-6 | 61.7 | 1.2 |
| S1 | ~550 | 72-82 | 17-19 | 7-9 | 34.4 | 3.8 |
| S2 | ~600 | 88-99 | 20-22 | 10-12 | 24.5 | 11.2 |
| S3 | ~650 | 105-116 | 23-25 | 13-15 | 10.2 | 13.7 |
| S4 | >700 | 121-210 | 26-42 | 16-32 | <5.7 | 70.1 |
*:样本量太小,资料缺如。(源自:Utermann,G.In:The metabolic andmolecular bases of inherited disease.7th ed, Vol.Ⅱ,eds.Scriver,C.R.etal.p1897,McGraw-Hill.Inc.,New York,1995)
由表4-6可看出,血浆Lp(a)的浓度亦与Kpn片段的大小成反比。这为血浆Lp(a)与Apo(a)多态型的大小成反比从基因DNA水平提供了直接的证据。
Scana等人在Kringle-4第37个重复区(KIV-37,又称为KringleⅣ-10)检测出了两种突变:一种突变导致第72个氨基酸被精氨酸取代;另一种突变导致第66个氨基酸苏氨酸被蛋氨酸取代。前者致使其Lp(a)不能与赖氨酸结合,且血浆Lp(a)浓度降低。后者似乎与Lp(a)浓度无关。Mooser等人报道在距第1第外显子5’端1.3Kb区或内,由5个核苷酸[(TTT-TA)n]构成的VNTR与其血浆Lp(a)浓度相关。此外。亦有报道Apo(a)基因5’端侧翼区DNA序列的多态性控制血浆Lp(a)的浓度。
四、ApoAⅠ基因的多态性ApoAⅠ为HDL的主要结构蛋白。ApoAI基因突变可导致异常的ApoAⅠ蛋白质的合成。有些异常的ApoAⅠ影响HDL的代谢。已报道的阻碍ApoAⅠ合成的基因突变有重排、缺失、无义突变等方式。这些突变的纯合子患者表现出极低HDL胆固醇水平和早发冠心病。目前已发现至少有20种不同的ApoAI结构基因的点突变导致氨基酸转换(见表4-7)。其中两种突变(精氨酸173→半胱氨酸和脯氨酸165→精氨酸)与低HDL胆固醇水平相关,但并不引致早发冠心病。
Talmud等人发现ApoAⅠ基因启动子区域内距转录起始部位第75个碱基(-75bp)A置换G的突变与高ApoAI及HDL水平相关。这种突变后的等位基因称为A等位基因,而含G的称为G等位基因。Saha等报道A等位基因在新加坡华人中出现的频率(0.27)高于高加索人(0.12)。Wang等人发现,ApoAⅠ基因第1内含子5‘末端,第83碱基C转换成T或第84碱基G转换成A,致使MspI酶切点丧失,这种突变与高HDL胆固醇相关,比上述的-75碱基A置换G突变与高HDL胆固醇的相关还强。
表4-7 人体ApoAI基因点突变导致氨基酸改变
| 氨基酸改变1 | 密码子2 | 核苷酸点突变 |
| Pro3→Arg | CCC | C→G |
| Pro3→His | CCC | C→A |
| Pro4→Arg | CCC | C→G |
| Arg10→Leu | CGA | G→T |
| Gly26→Arg | CGGC | G→C |
| Asp89→Glu | GATC | T→G,T→A |
| Asp103→Asn | CGAC | G→A |
| Lys107→0 | ||
| Lys107→Met | AAG | A→T |
| Glu136→Lys | CGAG | G→A |
| Glu139→Gly | GAG | A→G |
| Pro143→Arg | CCA | C→G |
| Glu147→Val | GAG | A→T |
| Ala158→Glu | GCG | C→G |
| Pro165→Arg | CCC | C→G |
| Glu169→Gln | CGAG | G→C |
| Arg173→Cys | GCGC | C→T |
| Arg177→His | CGC | G→A |
| Glu198→Lys | CGAG | G→A |
| Asp213→Gly | GAC | A→G |
1:氨基酸代号可参见表4-5脚注;第1排氨基酸右上角的数值表示其在ApoAⅠ一级结构上的位置;2:加底线的核苷酸为点突变部位。(源自:Eckardstein et al.J.Biol Chem.265:8612,1990)
五、其他载脂蛋白基因的多态性与以上介绍的ApoB、ApoE、Apo(a)以及ApoAI的基因一样,其他载脂蛋白的基因也存在多态性,以下简要地列举一些例子。
1.ApoAⅠ-CⅢ-AⅣ基因限制性片段长度多态性
在ApoAⅠ-CⅢ-AⅣ基因丛内,使用XmnⅠ、ApaⅠ、MspⅠ、PstⅠ、SstⅠ、XbaⅠ、TaqⅠ和PvuⅡ限制性内切酶已检测出10多种RFLP。尽管这些多态性大多由编码氨基酸以外的DNA序列突变所致,其中有些RFLP的携带者与增加冠心病的危险相关。比如用限制性内切酶PstⅠ消化从人群中搜集的DNA后。用ApoAⅠcDNA探针印迹DNA可测得3.3Kb的DNA片段(称为P2等位基因)与2.2Kb的DNA片段(称为P1等位基因)。P1等位基因在正常人群中出现率较高。P2等位基因在低HDL胆固醇患者中的出现率高于对照组的6倍。来自英国与美国的报道亦证实,杂合子P1P2携带者血浆ApoAⅠ与HDL胆固醇的水平显著地低于P1P2基因型携带者。这提示在ApoAⅠ-CⅢ-AⅣ基因丛中,PstⅠ酶切点的多态性,P2等位基因,与降低血浆ApoAⅠ和HDL的水平以及诱发冠心病有关。最近,Dallinga-Thie等人在美国脂类研究杂志上报道,用限制性内切酶XmnⅠ与MspⅠ在ApoAⅠ-CⅢ-AⅣ基因丛内检测出的稀有等位基因与高血浆胆固醇、甘油三酯、ApoB以及LDL胆固醇水平相连。这些结果显示,ApoAⅠ-CⅢ-AⅣ基因丛参与调节血浆胆固醇及甘油三酯的代谢。
2.ApoEⅠ-CⅠ-CⅡ基因限制性片段长度多态性
位于第19对染色体上的ApoEⅠ-CⅠ-CⅡ基因丛也有多态性的存在。检测出其RFLP的限制性内切酶包括:HpaⅠ、TaqⅠ、BglⅡ、DraⅠ、NcoⅠ以及BglⅡ。Klasen等人报道,HpaⅠ限制性片段长度的多态性与易发Ⅲ型高脂血症有关。然而HpaⅠ限制性片段长度的多态性与E2/E2基因型无关接关联。这说明HpaⅠ的RFLP是不同于ApoE基因的一种独立的遗传因子,其突变点尚未阐明。
3.ApoAⅡ基因的突变
Deeb等报道了在一对日本姐妹中检测出的家族性ApoAⅡ缺陷症。冠状动脉造影显示这对姐妹未患血管疾病。尽管免疫生化检验测不出血浆ApoAⅡ水平,患者HDL胆固醇在正常范围。DNA分析显示,患者ApoAⅡ基因内第3内含子剪接供体部位的第一个碱基G突变成为A,从而阻碍了内含子从初级转录物中剪除掉的过程。这无疑是导致ApoAⅡ缺陷的原因。
4.ApoAⅣ基因的多态性
常见的ApoAⅣ等位基因编码ApoAⅣ的第360与347个氨基酸,分别为谷氨酰胺和苏氨酸。当ApoAⅣ基因的第360密码子上第3个碱基T置换G后,正常的第360氨基酸位置上的谷氨酰胺被代之为组氨酸,当第347密码子上第1个碱基T置换A后,苏氨酸347→丝氨酸的变异即发生。关于这两种ApoAⅣ基因多态性对脂类代谢的影响尚无一致的结论。另一种突变,即密码子127的突变可致HincⅡ酶切位点的丧失。最近Kamboh等人对西伯利亚中部的驯鹿牧民检测中发现,没有HincⅡ酶切位点等位基因的携带者血浆甘油三酯水平高于HincⅡ酶切位点等位基因的携带者。
5.ApoCⅡ基因的多态性
载脂蛋白CⅡ基因的突变有四种类型被鉴定:a.无义突变;b.起动密码子的碱基突变;c.第2内含子剪切供体部位突变;d.由碱基插入或缺失所致的移码突变。这些突变患者的血浆ApoCⅡ水平显著降低或缺失。其遗传方式为常染色体隐性遗传。
6.ApoCⅢ基因的多态性
已发现有三种ApoCⅢ基因多态性与高甘油三酯血症有关:a.3175核苷酸C颠换成G;b.3206核苷酸T颠换成G;c.3206G等位基因。Li等人不久前在临床研究杂志上报道,ApoCⅢ基因启动子内-482与-455碱基的突变与发生高甘油三酯血症有关。作者推测这种突变正发生在受胰岛素抑制DNA序列范围内。突变后的ApoCⅢ基因启动子摆脱了胰岛素的抑制调控,ApoCⅢ的转录大大加强,ApoCⅢ的合成增加。这可能是人群中发生高甘油三酯的一个主要因素。
7.ApoD基因的多态性
Vijayaraghavan等人检测了57个肥胖病人57名对照者ApoD基因TaqⅠ酶切位点限制性片段长度多态性:2.2与2.7Kb等位基因。他们发现2.2Kb等位基因在肥胖者中的分布显著地高于正常者。这包括含有2.2Kb等位基因的杂合子与纯合子,提示显性遗传方式。作者认为,ApoD基因TaqⅠ限制性片段长度多态性可作为研究肥胖病的一个遗传标志。
8.ApoH基因的多态性
ApoH基因第247密码子突变,致使第247个氨基酸缬氨酸被亮氨酸取代。因为此突变使RasⅠ酶切位点丧失,故这种限制性片段长度多态性很易检测。最近Kamboh等调查西伯利亚中部驯鹿牧民中三种ApoH等位基因ApoH1、ApoH2、与ApoH3的分布与血浆脂质水平的关系,他们发现ApoH等位基因与男子低甘油三酯水平相关。而与女子高甘油三酯水平相关。而女子高甘油三酯水平相关。ApoH基因的多态性与脂质代谢的关系还有待于更广泛地、更深入地探讨。
9.ApoJ基因的多态性
Kamboh暨同事报道,在对美国白种人、黑人、美洲印第安人、爱斯基摩人、新几内亚人及尼日利亚人6个民族共985名受试者的检测中。他们发现了ApoJ有三种多态型,即ApoJ1、ApoJ2和ApoJ3。ApoJ1存在于所有受试者基因内。ApoJ2仅存于美国黑人与尼日利亚人黑人,出现频率分别为24%与28%。ApoJ3仅从一美国黑人受试者测出。造成此多态性的突变尚未见报道。这三种“等位基因”似乎对总胆固醇、LDL胆固醇、HDL胆固醇、VLDL胆固醇、甘油三酯的水平没有显著的影响。
结论随着分子生物学技术应用于脂类代谢与动脉粥样硬化的研究领域,几乎所有被命名的载脂蛋白的cDNA与基因被分离与鉴定。有关载脂蛋白基因多态性的报道与日俱增。人们对脂蛋白代谢紊乱从分子缺陷的水平进行认识取得了长足的进展。然而,各种载脂蛋白的生理功用尚未完全阐明。近年来发现ApoE4与老年性痴呆有关,揭示了ApoE的新的功用就佐证了这一点。对各种载脂蛋白的基因的各种多态性在所有种族间的分布进行系统的检测与分析还有待于进一步努力。一些新的、简易的、准确的检测方法也有待于开发。这些研究可望为载脂蛋白与动脉粥样硬化发病机制的探讨以及对动脉粥样硬化与冠心病的防治寻求新的措施开拓新的前景。
(叶永清)