第二节 抗原特异性基础
抗原的最大之一是其免疫效应具有特异性(specificity),这种特异性在其免疫原性和反应原性两方面都表现得非常突出。例如伤寒杆菌诱导的免疫应答只能针对伤寒杆菌;志贺杆菌不能诱导出对伤寒杆菌的免疫力,与抗伤寒杆菌抗体也不发生反应。这就是传统免疫学进行免疫预防和免疫诊断的基本依据。
抗原的特异性与蛋白分子中的氨基酸种类、排列顺序、特殊基因和空间构形等因素有关,甚至与其电荷性质及亲水性也有关系。但是其特异性不是平均地决定于整个分子,而是取决于分子表面几个氨基酸残基组成的特殊序列及其空间结构,称为表位(epitope)或抗原决定簇(antigenicdeterminant)。正是这些表位被淋巴细胞识别而诱导免疫应答,被抗体分子识别而发生抗原-抗体反应,这是研究抗原特异性的基础。
一、半抗原与载体(一)半抗原及其应用
免疫学先驱Landsteiner在本世纪初就已发现:某些不具有免疫原性的小分子物质可以与抗体结合,如果将其结合到具有免疫原性的大分子蛋白上就可诱导针对小分子物质的抗体应答。他借用希腊语“haptien”(原意为强加、抓牢)称这种小分子为hapten,汉语译为半抗原;而将半抗原赖以附着的蛋白质分子称为载体(carrier)。
结合到大分子载体上以后,半抗原可以改变载体原有的表位,也可以形成新的表位。半抗原在表位中是关键性的基团。抗原特异性的研究多是通过着抗原-载体复合物来进行的。
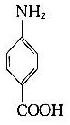
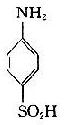
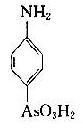
将苯胺、对氨基苯甲酸、对氨基苯磺酸和对氨基苯砷酸等四种已知结构的半抗原分别以偶氮方式结合到大分子蛋白上,然后用这些结合物分别免疫动物,结果所产生的抗体能够精确地将上述结构相似的半抗原区别开来,四种抗体只与相应的抗原发生反应(表1-1)。即使应用结构稍有差别的同一种物质,例如对位、邻位和间位的三种氨基苯甲酸作为半抗原,所产生的抗体也能将这些分子分出来。利用这种方法不仅可制备出针对许多小分子物质的抗体,甚至抗金属离子的抗体,而且大大地促进了对抗原性质的研究。
表1-1不同基团对抗原特异性的影响
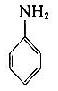 |  |  |  | |
| 抗苯胺 | + | - | - | - |
| 抗对氨基苯甲酸 | - | + | - | - |
| 抗对氨基苯磺酸 | - | - | + | - |
| 抗对氨基苯砷酸 | - | - | - | + |
(二)半抗原-载体效应
在半抗原-载体复合物中,载体分子虽有它本身的特异性,却不干扰半抗原的特异性。但是载体特异性对半抗原诱导抗体应答的效果有明显的影响。
将半抗原2,4-二硝基酚(DNP)共价交联于牛血蛋白(BSA)和卵白蛋白(OVA)等大分子载体上,对不同组的动物用不同的抗原进行首次和再次免疫,然后测定各组动物的抗DNP抗体,所得结果见表1-2。
表1-2半抗原-载体反应
| 首次免疫 | 再次免疫 | 抗DNP抗体 |
| DNP | - | |
| OVA | - | |
| OVA-DNP | + | |
| OVA-DNP | OVA-DNP | ++++ |
| OVA-DNP | BSA-DNP | + |
| OVA | OVA-DNP | ++++ |
| OVA | BSA-DNP | + |
从以上实验可以看出:载体不仅赋于半抗原以免疫原性,还与半抗原免疫应答的记忆性密切相关。进一步的研究证明,半抗原的特异性被B细胞识别,而载体特异性被T细胞识别;只有T-B细胞协作,才能启动对半抗原的抗体应答,才能产生再次应答效应。
二、天然抗原的表位对天然表位的研究不象半抗原-载体复合物那样简单,但是借助现代科学技术的进步,目前对表位已经有比较深刻的认识。
(一)表位的构成
表位只是抗原分子中几个氨基酸残基组成的特殊结构,在免疫效应中能全方位地与淋巴细胞或抗分子接触。抗体分子的抗原结合点并不很大,所以表位一般只占有大约3nm×1.5nm×0.7nm的空间,即5~7个氨基酸和单糖残基的大小,至多不超过20个氨基酸残基。
表位的构成方式至少有两种:①由某些氨基酸残基按一定顺序连续排列组成的线状序列,称为顺序(sequential)表位或线性(linear)表位(图1-1)。顺序表位是蛋白分子的一级结构,比较稳定,不受蛋白质加热变性和空间构形改变的影响。②由分子内不连续的2~3个氨基酸残基折叠排列所形成的三维结构构成,称为构象(conformational)表位(图1-1);有时候,呈α螺旋式排列的连续肽链序列也可起到构象表位的作用。构象表位的抗体可用来研究蛋白分子在生理或病理过程中三维结构的变化。但是构象表位是蛋白质的二级或三级结构,不太稳定,在蛋白质受热或酶解变性后会彻底破坏,不能恢复。因此分离和研究比较困难。

图1-1顺序表位和构象表位示意图
(二)表位的数目和定位
抗原分子上表位的数目可以用饱和情况下能够结合多少个抗体分子来测定,一般情况下表位数目与抗原的分子量呈正相关。例如鸡卵蛋白的分子量为42kD,有5个表位;甲状腺球蛋白的分子量700kD,有大约40个表位。一个表位能结合抗体分子上的一个抗原结合点,所以可将抗原分子表位的数目称为抗原的结合价。例如上述鸡卵蛋白为5价,甲状腺球蛋白为40价。
虽然一个抗原分子上可以有多个表位,但在诱导宿主免疫应答时可能有一种或一个表位起主要作用,使宿主产生以该特异性为主的免疫应答;这种现象称为免疫显性或免疫优势(immunodominance);起关键作用的表位称为显性表位。这个原则也适用于一个表位中不同的氨基酸残基,在表位中也有所谓的显性基团存在,如被置换会明显改变表位特异性。这种现象可能与表位在抗原分子中的位置或显性基团在表位中的位置有关。上述半抗原就是在表位中起到显性基团的作用。
实验证明,表位只有位于抗原分子的表面才能与淋巴细胞和抗体分子接触而发挥免疫效应。用多聚赖氨酸为骨架,用丙氨酸(A)、谷氨酸(G)和酪氨酸(T)序列为支链进行的研究能清楚地证明这一点:将G和T连在骨架外部,使可诱导以针对G和T为优势的抗体产生;将连续的A连在骨架外部,便诱导以针对A为优势的抗体产生。
(三)共同表位与交叉反应
一个抗原分子上可能只有一种表位,称为单纯抗原;但是天然情况下很少发现单纯抗原,多数抗原分子上都存在多种表位。一般地说,不同的抗原物质具不同的表位,故各具特异性;但有时某一表位也会出现在不同的抗原上,称为共同表位,带有共同表位的抗原互称共同抗原。拥有共同抗原在自然界、尤其在微生物中是很常见的一种现象,存在于同一种属或近缘种属中称为类属抗原,存在于远缘不同种属中则称为异嗜性抗原(heterophileantigen)。
共同表位的例子很多,例如沙门菌可根据其O抗原分为40多个血清组,含2000多个血清型,同一组成员都有共同的O表位,是由特定的单糖决定的。再如人类、动物、植物和微生物之间也广泛存在着一种以发现者名字命名的抗原,称为Forssman抗原,其共同表位由共价交联于神经酰胺脂上的N-乙酰半乳糖胺、半乳糖和葡萄糖联合组成。有些共同表位只是结构相似,所以又称相似表位。
由某一抗原诱导产生的抗体,也可以与其共同抗原结合,这种现象称为交叉反应(crossreaction)。这种交叉反应可用来解释某些免疫病理现象,也可以用来诊断某些传染病。但是交叉反应总不如抗体与其诱导抗原之间的结合那么牢固;在与交叉抗体结合时只能部分地吻合,不能达到整个空间的完全相配。
(四)表位的细胞识别性
抗原免疫原性的体现首先是淋巴细胞对表位的识别。有证据表明,细胞介导免疫和体液免疫是针对同一抗原分子的不同部分,例如用人的高血糖素对小鼠进行免疫,产生的抗体是针对其氨基酸,而细胞介导免疫则是针对其羧基端;结论是T细胞和B细胞识别的不是同一类表位。这样可以将表位分为两类:B细胞(识别)表位和T细胞(识别)表位。
1.B细胞表位供B细胞识别诱导抗体应答,而T细胞不能识别的表位。对这类表位的研究资料较多,以上所述几乎都是应用特异性抗体、针对B细胞表位进行研究而得出的,此处不再赘述。
2.T细胞表位供T细胞识别诱导产生细胞介导免疫,而B细胞不能识别的表位。T细胞的抗原受体露出膜外部分较少,不能象抗体分子那样结合游离的抗原,只能识别由抗原递呈细胞(见第四章)递呈的与MHC结合的表位。所以被T细胞识别的抗原必须事先经过一定的处理,从蛋白质降解为多肽,再与MHC分子结合。因为构象表位在蛋白质降解时会遭到破坏,无法被T细胞识别,所以T细胞表位主要是顺序表位,也不一定位于分子表面。象抗体分子一样,T细胞也可以由共同抗原引起交叉反应,但总不如与原诱导抗原的结合那么有效。
T细胞表位可以诱导细胞免疫应答,作为细胞毒性T细胞攻击的靶子,同时对诱导抗体应答也是必需的;因为B细胞的活化需要活化T细胞的辅助,而T细胞活化必须由T细胞表位来启动。由此看来,每个抗原分子必须至少有一个T细胞表位,才能使抗原具有免疫原性。只具有B细胞表位的分子可以作为抗体的靶子,但本身不能诱导抗体应答。只有少数分子可能例外。