第三节 MHC分子
虽然不同个体、不同种属的MHC结构不同,但其编码的分子在化学结构、组织分布及功能上均十分相近,可以分成三类:即Ⅰ类分子,Ⅱ类分子和Ⅲ类分子;其编码基因也相应地分成三类。Ⅰ类和Ⅱ类分子是结构相似的细胞膜表面糖蛋白,除作为移植抗原外,还与抗原递呈及某些疾病相关。Ⅲ类分子包括C2、C4、B因子和肿瘤坏死因子等多种可溶性蛋白质,相互间差别很大,与Ⅰ类和Ⅱ类分子在结构和功能上相关性很小,本章不予讨论。
一、MHCⅠ类分子(一)Ⅰ类分子的结构
人类的类分子由HLA的A、B、C、E、F、G、H、K和L等基因编码;但因后几类基因的性质和作用尚不清楚,所以目前所称的类分子主要指HLA-A、B、C位点的抗原。
Ⅰ类分子是由非共价键连接的两条肽链组成的糖蛋白;其中一条称为重链或α链,另一条为轻链或称为β2微球蛋白(β2m)。α链的分子量为44kD,结构呈多态性;其羧基端穿过细胞膜,伸入胞浆之中,氨基端则游离于细胞膜外(图6-3)。α链的膜外区肽段折叠形成三个功能区,分别称为α1、α2、和α3区;每个功能区约含90个氨基酸残基,其结构与Ig相似;α1和α2区的氨基酸顺序变化较大,决定着Ⅰ类分子的多态性。β2m不MHC基因编码,而是第15号染色体上单个基因编码的产物,分子量12kD。其结构与Ig恒定区(CH3)有较大同源性,属于Ig超族成员,没有同种异型决定簇,但具有种属特异性。β2m不穿过细胞膜,也不与细胞膜接触,而是以非共价形式附着于α3的功能区上。虽然β2m不直接参与Ⅰ类分子的抗原递呈过程,但是它能促进内质网中新合成的Ⅰ类分子向细胞表面运输,并对稳定Ⅰ类分子的结构具有一定作用。
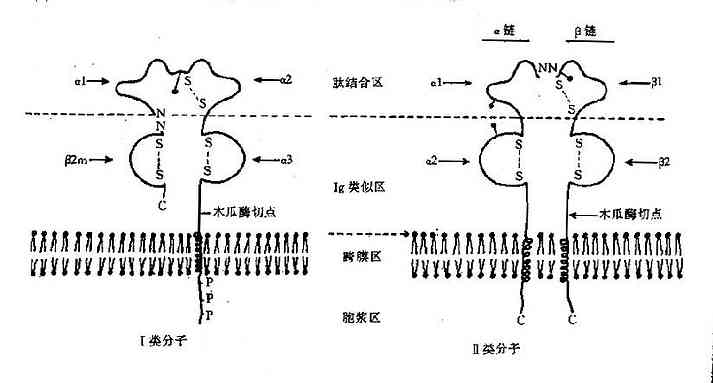
图6-3HLA分子结构示意图
利用X线结晶衍射图分析,阐明了Ⅰ类分子的三维结构:α1和α2功能区共同构成了槽形的抗原肽段结合部位,来自α1和α2的8条反向平行的β片层结构组成槽的底部;而α1和α2功能区的其余部分各自形成一个α-螺旋,两条螺旋相互毗邻,相互平行,在分子的远端共同构成槽的侧壁;槽的底部和侧壁体现Ⅰ类分子的多肽性,也决定了Ⅰ类分子与抗原结合的特异性;α3功能区具有与CD8分子结合的空间构型(图6-4)。
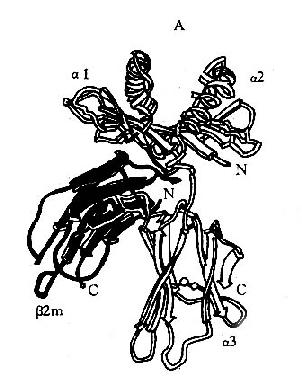
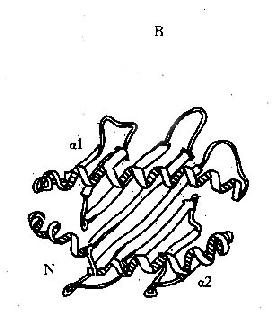
图6-4HLAⅠ类分子多肽折迭立体结构示意图
A:侧面观;B:上面观
Ⅰ类分子与抗原的结合有一定的选择性,但是没有抗体和TCR与抗原结合的特异性高。Ⅰ类分子的抗原结合槽能够容纳8~10个氨基酸长度的抗原片段,其抗原结合点主要与氨基酸顺序相对恒定的抗原肽段的骨干结合,而将抗原肽段上多变的氨基酸侧链处于游离状态;这些侧链却能与TCR和抗体结合。每个细胞表面可以表达约106个Ⅰ类分子,每个Ⅰ类分子都能与相当各类的抗原肽段结合。因此每个细胞都具有同时递呈许多不同抗原的潜力,从而保证一个正常的个体对绝大多数抗原发生Ⅰ类限制性的免疫应答。
(二)Ⅰ类分子的分布和功能
MHCⅠ类分子分布于几乎所有有核细胞表面,但不同组织细胞的表达水平差异很大:淋巴细胞表面Ⅰ类抗原的密度最高,肾、肝、肺、心及皮肤次之,肌肉、神经组织和内分泌细胞上抗原最少,而成熟红细胞、胎盘滋养层细胞上未能检出,血清、尿液中及初乳等体液中也有可溶性形式存在的Ⅰ类抗原。干扰素、肿瘤坏死因子在体内外均可增强各种细胞对Ⅰ类分子的表达。
Ⅰ类分子的重要生理功能是对CD8+T细胞的抗原识别功能起限制性作用,也就是参与向CD8+T细胞递呈抗原的过程。CD8+T细胞只能识别与相同Ⅰ类分子结合的抗原(多为内源性的细胞抗原,如病毒感染的细胞和肿瘤细胞等),这种现象称为MHC限制性。例如,当病毒感染了某个细胞,病毒抗原可被分解成一些短肽片段,后者与在内质网中合成的Ⅰ类分子结合后表达于细胞表面,才能被CD8+T细胞识别。Ⅰ类分子主要介导Tc细胞的细胞毒作用,也是重要的移植抗原。
二、MHCⅡ类分子(一)Ⅱ类分子的结构
人类的MHCⅡ类分子由HLA复合体中的D区基因编码,已经明确的Ⅱ类分子包括HLA-DR、DP和DQ抗原。Ⅱ类分子亦是由非共价连接的两条多肽链组成,分别称为α链和β链;与Ⅰ类分子不同的是,两条链均由HLA基因编码。α链的分子量约34kD,β链约29kD;两条肽链均嵌入细胞膜,伸入胞质之中;其膜外区各有两个Ig样的功能区(图6-3),分别称为α1、α2、β1和β2功能区。
X线结晶衍射图显示,Ⅱ类分子的α1和β1功能区共同形成一个与Ⅰ类分子相似的槽型结构的多肽结合区。1和β1各有一个螺旋,形成槽的两侧壁,其余部分形成片层,构成槽的底部。Ⅱ类分子的多态性也体现在多肽结合槽的侧壁和底部,所以其空间构型依编码基因的不同而异。类分子的抗原结合特性亦与Ⅰ类分子一样,特异性不强,每个分子可能与多种肽片结合;但与Ⅰ类分子不同的是,Ⅱ类分子肽结合槽的两端呈开放状(Ⅰ类分子的结合槽两端呈封闭状),能够容纳较长(10~18个氨基酸残基)的肽段。
(二)Ⅱ类分子的分布和功能
Ⅱ类分子的分布比较局限,主要表达于B细胞、单核-巨噬细胞和树突状细胞等抗原递呈细胞上,精子细胞和某些活化的T细胞上也有Ⅱ类分子。一些在正常情况下不表达Ⅱ类分子的细胞,在免疫应答过程中亦可受细胞因子的诱导表达Ⅱ类分子,因此Ⅱ类分子的表达被看成是抗原递呈能力的标志。IL-1、IL-2和干扰素在体内外均能增强Ⅱ类分子的表达。有些组织在病理条件下也可表达一些类抗原,如胰岛β细胞、甲状腺细胞等。
Ⅱ类分子的功能主要是在免疫应答的始动阶段将经过处理的抗原片段递呈给CD4+T细胞。正如CD8+T细胞只能识别与MHCⅠ类分子结合的抗原片段一样,CD4+T细胞只能识别Ⅱ类分子结合的抗原片段。Ⅱ类分子主要参与外源性抗原的递呈,在一些条件下也可递内源性抗原。在组织或器官移植过程中,Ⅱ类分子是引起移植排斥反应的重要靶抗原,包括引起宿主抗移植物反应(HVGR)和移植物抗宿主反应(GVHR)。在免疫应答中,Ⅱ类抗原主要是协调免疫细胞间的相互作用,调控体液免疫和细胞免疫应答。