第四节 脱氧核糖核苷酸的生成
DNA与RNA有两方面不同:(1)其核苷酸中戊糖为2?脱氧核糖而非核糖。(2)含有胸腺嘧啶碱基,不含尿嘧啶碱基。
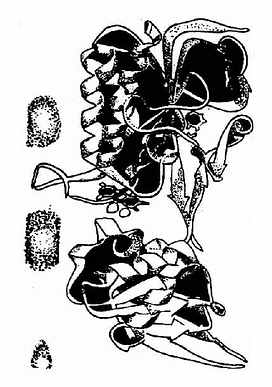
图8-11 大肠杆菌硫氧化还原
蛋白的320残基亚单位结构图
(一)脱氧核糖的生成:脱氧核糖核苷酸是通过相应核糖核苷酸还原,以H取代其核糖分子中C2上的羟基而生成,而非从脱氧核糖从头合成。此还原作用是在二磷酸核苷酸(NDP)水平上进行的。(此处N代表A、G、U、C等碱基)。
催化脱氧核糖核苷酸生成的酶是核糖核苷酸还原酶(ribonudeotide reductase)。已发现有三种不同的核糖核苷酸还原酶,此反应过程较复杂。核糖核苷酸还原酶催化循环反应的最后一步是酶分子中的二硫键还原为具还原活性的巯基的酶再生过程。硫氧化还原蛋白(thioredoxin)是此酶的一种生理还原剂,由108个氨基酸组成,分子量约12kD。含有一对邻近的半胱氨酸残基(图8?11)。所含硫基在核糖核苷酸还原酶作用下氧化为二硫键,后者再在在硫氧化还原蛋白还原酶(thioredoxinreductase)催化,由NADPH供氢重新还原为还原型的硫氧化还原蛋白。因此,NADPH是NDP还原为dNDP的最终还原剂。(图8-12)
核糖核苷酸还原酶是一种变构酶,包括B1、B2两个亚基,只有B1与B2结合时。才具有酶活性。在DNA合成旺盛、分裂速度快的细胞中,核糖核苷酸还原酶系活性较强。
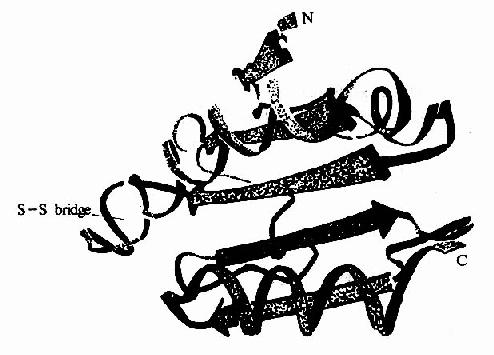
图8-12 大肠杆菌硫氧化还原蛋白的X线衍射结构
(二)脱氧核糖核苷酸合成的调节四种dNTP的合成水平受到反馈调节,同时保持dNTP的适当比例也是细胞正常生长所必需的。实际上,缺少任一种dNTP都是致命的,而一种dNTP过多也可致突变,因为过多的dNTP可错误掺入DNA链中。核糖核苷酸还原酶的活性对脱氧核糖核苷酸的水平起着决定作用。各种dNTP通过变构效应调节不同脱氧核糖核苷酸生成。因为,某一种特定NDP经还原酶作用生成dNDP时,需要特定NTP的促进,同时受到另一些NTP的抑制(表8?2)。通过调节使4种dNTP保持适当的比例。
表8-2 核糖核苷酸还原酶的别构调节
| 作用物 | 主要促进剂 | 主要抑制剂 |
| CDP | ATP | dATP、dGTP、dTTP |
| UDP | ATP | dATP、dGTP |
| ADP | dGTP | dATP、ATP |
| GDP | dTTP | dATP |
例如,当存在混合的NDP底物时,由ATP促使CDP和UDP还原生成dUDP和dCDP。经dUDP转变为dTTP(后述),dTTP则反馈抑制CDP和UDP还原,同时促进dGDP的生成,dGDP磷酸化生成dGTP则抑制GDP、CDP和UDP的还原,而促进ADP的还原生成dADP。当dATP升高与酶
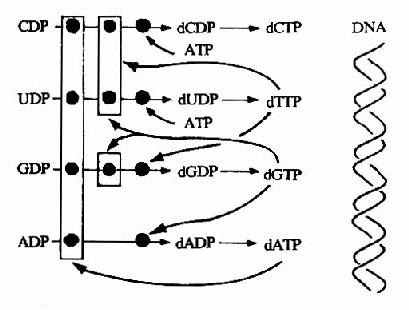
图8-13 脱氧核苷酸合成调节网络
活性位点结合,则抑制所有NDP的还原反应(图8-13)。细胞内dCTP和dTTP的适当比例并非由核糖核苷酸还原酶调节,而是通过脱氧胞嘧啶脱氨酶(deoxycytidinedeaminase)决定。此酶催化dUMP的生成,dUMP则是dTTP的前体。此酶受dCTP激活,受dTTP抑制。
dNTP由dNDP磷酸化生成:
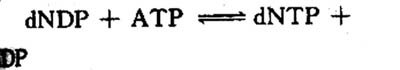
由二磷酸核苷酸激酶(mucleosidediphosphafe kinase),催化与催化NDP磷酸化的反应相似。
(三)脱氧胸腺嘧啶核苷酸的生成脱氧胸腺嘧啶核苷酸(dTMP)是由脱氧尿嘧啶核苷酸(dUMP)甲基化生成。而dUMP由dUTP水解生成:
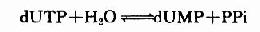
体内进行此种“浪废”能量的反应过程的意义在于:细胞必须减少细胞内dUTP浓度以防止脱氧尿嘧啶掺入DNA中,因为合成DNA的酶系不能有效识别dUTP和dTTP。
dUMP甲基化生成dTMP由胸腺嘧啶合成酶(thymidylatesynthetase,TS)催化,N5,N10-甲烯FH4提供甲基(图8-14)。N5、N10-甲烯-FH4提供甲基后生成的FH2又可以再经二氢叶酸还原酶的作,重新生成四氢叶酸。
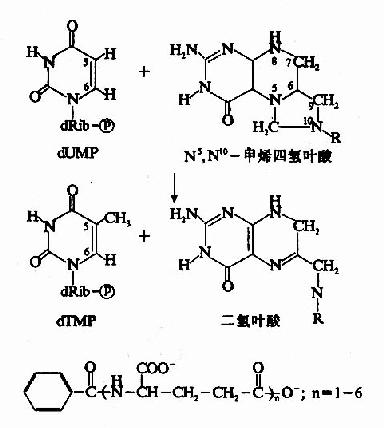
图8-14 dTMP的生成