第三节 神经递质(Neurotransmitter)的代谢
神经系统内存在着许多化学物质,但它们不一定都是递质(transmitter)。作为神经递质,必须具备几个基本条件:(1)被认为是递质的物质应特异性地存在于以该物质为递质的神经元中,而且,在这种神经元的末梢有合成该递质的酶系统;(2)递质在神经末梢内合成以后,通常是集中贮存在囊泡(vesicle)内,这样可以防止被胞浆内的其它酶所破坏;(3)当神经冲动到来时,神经末梢内的递质就自突触前膜释放入突触间隙;(4)递质通过突触间隙,作用于突触后膜的叫做受体的特殊部位,引起突触后膜离子通透性改变以及电位变化;(5)神经递质在发挥上述效应后,其作用应该迅速终止,以保证突触传递的高度灵活。作用的终止有几种方式:一是被酶所水解,失去活性;其次是被突触前膜“重摄取”,或是一部分为后膜所摄取;也有的部分进入血循环,在血中一部分被酶所降解破坏。
目前已知的神经递质种类很多,但主要的有乙酰胆硷、儿茶酚胺类(去甲肾上腺素和多巴胺)、5?羟色胺、GABA、某些氨基酸和寡肽等。
一、乙酰胆碱(Acetylcholine简写Ach)乙酰胆碱是许多外周神经如运动神经、植物性神经系统的节前纤维和付交感神经节后纤维的兴奋性神经递质。
(一)合成和贮存
Ach由胆碱和乙酰CoA所合成。胆碱乙酰化酶(choline acetylase)催化下列反应:
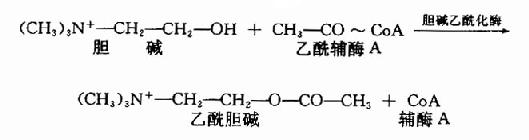
由于胆碱乙酰化酶位于胞浆内,因此设想Ach是先在胞浆内合成,然后进入囊泡贮存。平时囊泡中和胞浆中的Ach大约各占一半,且两者可能处于平衡状态。囊泡内贮存的Ach是一种结合型的(与蛋白质结合),而释放至胞浆时,则变为游离型。
(二)释放、重摄取和失活
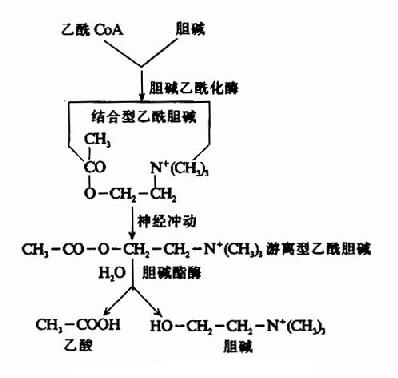
当神经冲动沿轴突到达末梢时,囊泡趋近突触膜,并与之融合、破裂,此时囊泡内结合型Ach转变为游离型Ach,释放入突触间隙。同时,还可能有一部分胞浆内新合成的Ach也随之释放。
Ach作用于突触后膜(突触后神经元或效应细胞的膜)表面的受体,引起生理效应。已经确定Ach受体是一种分子量为42,000的蛋白质,通常以脂蛋白的形式存在于膜上。
Ach在传递信息之后和受体分开,游离于突触间隙,其中极少部分在突触前膜的载体系统作用下重新被摄入突触前神经元。大部分Ach是在胆碱酯酶的作用下水解成胆碱和乙酸而失去活性,也有一部分经弥散而离开突触间隙。关于乙酰胆碱在神经末梢中的代谢,可总结如图(14-4,14-5)。
 |  |
| 图14-5 乙酰胆碱的代谢 | 图14-4 突触部位Ach的动态 |
儿茶酚胺类是指含有邻苯二酚基本结构的胺类。体内具有生物活性的儿茶酚胺包括多巴胺(dopamine,DA)、去甲肾上腺素(norepinephrine,noradrenaline,NE)和肾上腺素(epinephrine,adrenalin,E)。它们的结构如下。
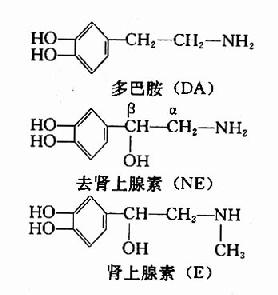
去甲肾上腺素和肾上腺素既是肾上腺髓质所分泌的激素,又是交感和中枢神经系统中去甲肾上腺素能纤维的神经递质。NE在中枢内分布广泛,含量较多,E则少,因此我们着重介绍NE的代谢。DA则主要集中在锥体外系,也是一种神经递质。
(一)儿茶酚胺的生物合成
神经组织中儿茶酚胺的合成原料来自血液中的酪氨酸,其合成过程如图14-6。
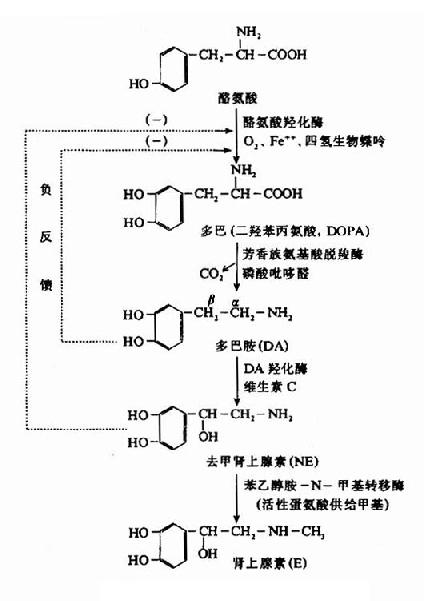
图14-6 儿茶酚胺类的生物合成
在上述过程中,第一步有酪氨酸羟化酶参加,它位于去甲肾上腺素能神经纤维的胞浆内,含量少,活性低,成为NE生成的限速酶,四氢生物蝶呤是它的辅酶,O2和Fe++也是合成时必不可少的因素;第二步反应是由胞浆中芳香族氨基酸脱羧酶所催化,这酶的特异性不高,和一般氨基酸脱羧酶一样,要求磷酸吡哆醛作为辅酶;第三步是多巴胺羟化酶催化的氧化反应,氧化发生在β碳原子上,此酶不存在于胞浆而是附于囊泡内壁,属于含Cu++的蛋白质,并需要维生素C为辅助因子。
从以上各酶的亚细胞水平分布可以了解,合成NE的最后一步只能在囊泡内进行,NE合成量不仅受酪氨酸羟化酶限速调节,而且当神经末梢胞浆中游离的NE浓度过高时,还可以因反馈性抑制酪氨酸羟化酶的作用而减少NE的合成。
苯乙醇胺-N-甲基转移酶主要见于肾上腺髓质细胞,可使NE甲基化生成肾上腺素。此酶在脑内虽有少量存在,但一般认为正常时哺乳类脑内的肾上腺素含量极少。有人认为,如果苯乙醇胺?N?甲基转移酶活性过高,可以使多巴胺直接变成N?甲基多巴胺等物,造成这些递质的代谢紊乱,这可能是精神分裂症的原因之一。
(二)去甲肾上腺素(NE)
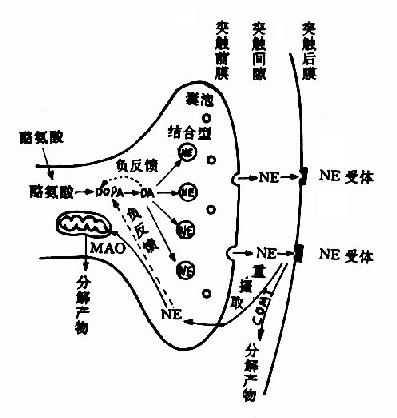
1.贮存和释放 NE在囊泡内合成后就地贮存。它和ATP、嗜铬颗粒蛋白等疏松地结合在一起,这样,就使NE不易渗入胞浆而遭单胺氧化酶的破坏。当神经冲动到达末梢时,突触前膜附近的囊泡便与前膜融合,破裂并生成小孔。这时囊泡内的NE连同嗜铬颗粒蛋白等一起被释放入突触间隙。
2.突触间隙中NE的去向 释放入突触间隙的NE能与突触后膜上的NE受体结合,产生生理效应。然后,约有3/4的NE重新被突触前膜所摄取。摄入胞浆后的NE又可进入囊泡贮存。重摄取是一种耗能过程,与突触前膜上的Na+,K+?ATP酶系统及囊泡膜上的Mg++-ATP酶系统有关。另外一部分NE则为后膜所摄取,并在那里被分解灭活。还有一部分是在突触间隙中被破坏或逸入血液。除了被突触前膜和囊泡重摄取的NE可再供利用外,其余的NE大都遭到酶促降解而灭活。NE的酶促降解如图14?所示,单胺氧化酶(MAO)和儿茶酚-O-甲基转移酶(catechol?o?transmethylase,COMT)是催化儿茶酚胺分解的两种主要的酶,它们不仅存在于神经组织内,而且广泛地分布于非神经组织,神经元的线粒体膜上也很多。NE受MAO作用,首先氧化脱氨基生成醛,后者再变成醇或酸。3-甲氧基-4-羟基苯乙二醇(MHPG)是中枢内NE的主要降解产物。在外周则以氧化成香草基扁桃酸(VMA)为主。血液循环中的NE(主要是激素),则在肝、肾等组织经COMT作用变成甲氧基代谢产物而排泄。现在,临床上常测定尿中VMA含量,作为了解交感神经功能的指标,患嗜铬细胞瘤和神经母细胞瘤时,由于肿瘤组织也产生NE或E,其代谢产物VMA相应增多,故在诊断上颇有意义。
关于NE在突触部位的动态可用图14-8表示。
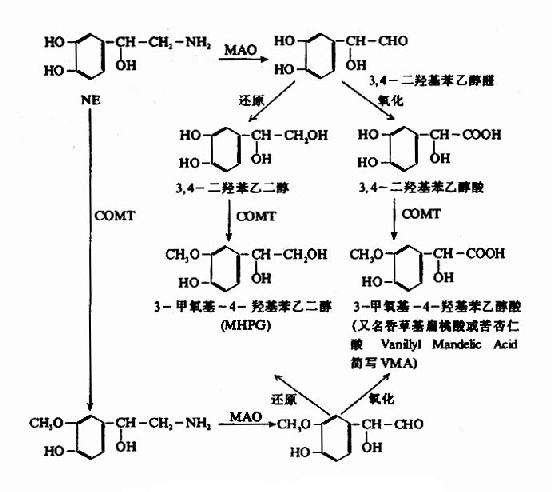
图14-7 去甲肾上腺素的分解代谢
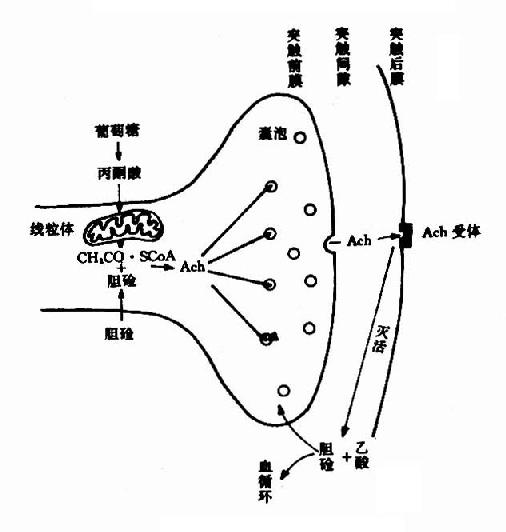
图14-8 突触部位NE的动态
中枢神经系统内的去甲肾上腺素能神经元胞体集中在延脑和桥脑,其通路都已研究明确。但是NE在中枢究竟是抑制性还是兴奋性的递质,目前尚不能肯定,可能随部位不同而异。NE所引起的生理效应也很难用简单的术语“兴奋”、“抑制”来表达。根据一些动物实验观察到,NE可引起动物嗜睡,体温降低,出现摄食行为。有人认为,脑内NE减少,可表现出精神抑郁;反之,过量可表现出狂躁。总之,脑内NE的功能可能和体温、摄食行为、镇痛、心血管系统和精神状态的调节有密切关系。
(三)多巴胺(DA)
在儿茶酚胺类递质的生物合成过程中,多巴胺是去甲肾上腺素的前体。体内凡有NE的组织,其中必然也有DA。由于DA在中枢某些部位中浓度很高,而且它的分布又与NE不平行,所以一般认为它本身也是一种独立的神经递质。
脑内多巴胺的作用是多方面的,它可能和躯体运动功能的加强,垂体内分泌机能的加强以及精神活动的调节都有关系。
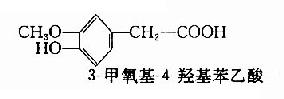
多巴胺能神经末梢中的囊泡是贮存DA的场所。这种囊泡不同于NE囊泡,前者不含多巴胺?β?羟化酶,所以不致将DA羟化成NE。此外,在去甲肾上腺素能纤维中,NE囊泡对贮存物的要求是β?位置上有羟基,而DA结构上没有β?羟基。DA的贮存、释放和酶促降解都和NE十分相似,而更新速度则较NE快。脑内DA的代谢产物主要是3-甲氧基-4-羟基苯乙酸(homovanillic acid简写HVA,又名同型香草酸)
5-羟色胺又名血清紧张素(serotonin),最早是从血清中发现的。中枢神经系统存在着5-?色胺能神经元,但在脊椎动物的外周神经系统中至今尚未发现有5-羟色胺能神经元。
由于5-羟色胺不能透过血脑屏障,所以中枢的5-羟色胺是脑内合成的,与外周的5-羟色胺不是一个来源。用组织化学的方法证明,5-羟色胺能神经元的胞体在脑内的分布主要集中脑干的中缝核群,其末梢则广泛分布在脑和脊髓中。
(一)合成、贮存和释放
5-羟色胺的前体是色氨酸。色氨酸经两步酶促反应,即羟化和脱羧,生成5-羟色胺。此过程在某种程度上和儿茶酚胺的生成相似。
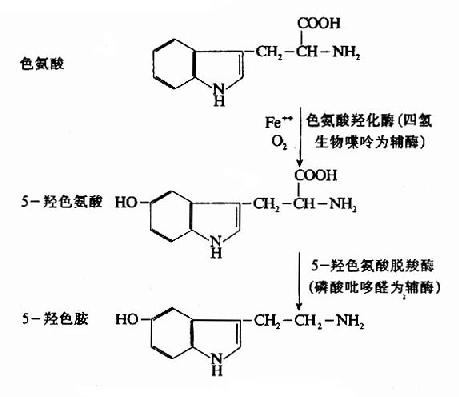
图14-9 5-HT的生成
色氨酸羟化酶象酪氨酸羟化酶一样,需要O2、Fe++以及辅酶四氢生物蝶呤。但脑内这种酶的含量较少,活性较低,所以它是5-HT生物合成的限速酶。此外,脑内5-HT的浓度影响色氨酸羟化酶的活性,从而对5-HT起着反馈性自我调节作用。血中游离色氨酸的浓度也影响脑内5-HT的合成,当血清游离色氨酸增多时(例如给大鼠腹腔注射色氨酸后),进入脑的色氨酸就增多,从而加速了5-HT的合成。
(二)重摄取和降解
和儿茶酚胺类递质一样,释放到突触间隙的5?HT,大部分被突触前神经末梢重摄取,而且重摄取后,部分进入囊泡再贮存,部分则被线粒体膜上的单胺氧化酶(MAO)所氧化:
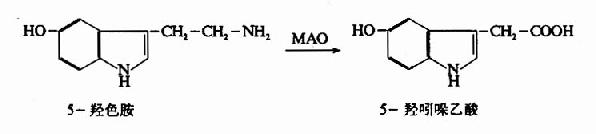
这就是脑内5-HT降解的主要方式,5-羟吲哚乙酸(5-hydroxyindolacetic acid)无生物活性。
检查5-HT对各种神经元的作用时发现,5-HT可使大多数交感节前神经元兴奋,而使付交感节前神经元抑制。损毁动物的中缝核或用药物阻断5-HT合成,都可使脑内5-HT含量明显降低,并引起动物睡眠障碍,痛阈降低,同时,吗啡的镇痛作用也减弱或消失。如果电刺激大鼠的中缝核,可影响其体温升高;另一方面,也观察到室温升高时大鼠脑内5-HT更新加速。这些现象揭示脑内5-HT与睡眠、镇痛、体温调节都有关系。还有人报导,5-HT能改变垂体的内分泌机能。此外,有人提出5-HT能神经元的破坏是精神性疾病时出现幻觉的原因。可见精神活动也与5?HT有一定的关系。
四、氨基酸和寡肽脑内到处都存在着氨基酸。过去只认为它们是合成蛋白质的原料,或是蛋白质分解的产物。近年来,注意到某些氨基酸在中枢的突触传递中起着递质的作用。而且发现,凡是中性氨基酸,如γ?氨基丁酸、甘氨酸、β?丙氨酸等对中枢神经元表现抑制作用,而酸性氨基酸如谷氨酸、天门冬氨酸则表现为兴奋作用。
有一些小分子肽类在中枢神经系统中也具有神经递质同的作用。1975年发现的脑啡肽(enkephalin)是由五个氨基酸残基构成的寡肽,从猪脑中分离出来的脑啡肽又分为两型。
蛋氨酸型(蛋脑啡肽,Met-enkephalin):
H-酪-甘-甘-苯丙-蛋-OH
亮氨酸型(亮脑啡肽,Leu-enkephalin)
H-酪-甘-甘-苯丙-蛋-OH
它们是由脑细胞内合成的具有吗啡样作用的肽,故名脑啡肽。已知吗啡(morphine)要先与脑内的吗啡受体结合才能产生镇痛、欣快作用,而脑啡肽之所以有吗啡样作用也是因为它们能与吗啡受体结合。吗啡是外来的物质,而脑啡肽是内源性的。此外,已知脑内还有一些能与吗啡受体结合并产生吗啡样作用的其它的肽类,称之为内啡肽(endorphin),虽然还不能十分肯定这些肽类都是真正的神经递质(即完全符合前述神经递质的条件),但是,研究它们的作用对阐明脑的功能,特别对阐明痛觉原理是很有意义的。